social contact isn’t “the same stimulus” for every nervous system.
For a lot of people, being with others steadily becomes stabilizing. For me, it steadily becomes load-bearing. Not because I “don’t like people,” and not because I’m failing at connection—but because my system changes modes the moment another person is present.
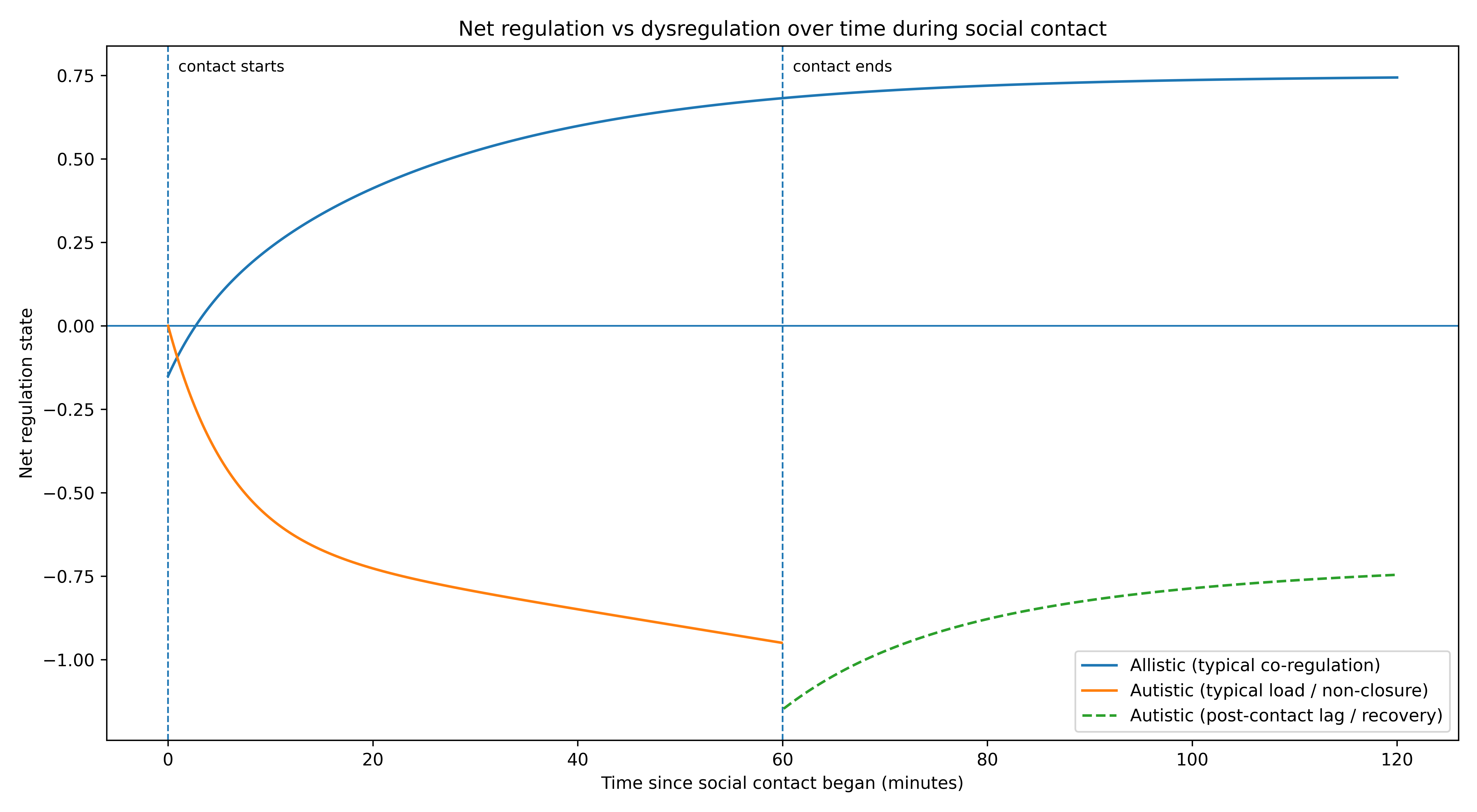
How to read it
The x-axis is time after social contact begins.
The y-axis is net state relative to baseline (0).
0 isn’t “perfectly regulated.” It’s a reference point: the equilibrium I can reach in solitude—where my perception settles, my feelings attach to the present, and my responses aren’t pre-audited against another mind.
Two trajectories
The upward curve (allistic co-regulation) represents a pattern many people describe intuitively: initial orientation cost, then increasing ease. The longer the interaction remains safe, the more the body treats it as resource—shared attention, shared context, reduced vigilance.
The downward curve (autistic coupling cost) represents something qualitatively different: the longer the interaction continues, the more unresolved computation accumulates—interpretation branches, consequence simulation, self-monitoring, sensory load, timing pressure. Even when I trust the person. Even when nothing “bad” is happening. The mere fact of being mutually present seems to keep my system in an interactive mode that resists closure.
And that’s the part that matters most to me:
closure timing is agency.
If my internal signal becomes coherent only after the moment has passed, then real-time influence becomes structurally asymmetric. I’m forced to choose between acting before I have my authentic read (which feels like betrayal) or waiting for the authentic read and losing my chance to shape what’s happening. That tradeoff isn’t a personality quirk. It’s a timing mismatch between inner convergence and outer pacing.
Why the dashed tail exists
The dashed segment after “contact ends” is there because the cost doesn’t necessarily stop when the interaction stops. Sometimes the body only releases enough for integration once the coupling variable goes to zero. So the “real feeling” arrives late—not as a surprise, but as the system finally finishing its work.
A note on “missions”
This is why a clear near-term objective can feel like oxygen. A defined objective reduces interpretive branching and compresses social ambiguity into a single constraint: “this is what we’re doing.” That doesn’t make the social state effortless—it just makes it computable in time.
And there’s a dark twin to that relief: the same mechanism is exploitable. If “mission clarity + permission” is what restores your agency in the moment, then anyone who can supply crisp objectives (especially with conditional approval) can quietly reroute your authorship—because saying yes feels like regulation, while resisting feels like returning to the ambiguity-paralysis you were trying to escape.
The main point
When people talk past each other about socializing—“it restores me” vs “it scrambles me”—they’re often describing two different regulation dynamics, not two different attitudes.
One system gets more stable through contact.
Another system pays a compounding coupling tax through contact.
Same room. Same conversation. Different bodily economics.
And once you see it that way, a lot of moralizing dissolves. What’s left is mechanics—and the relief of finally having a clean model for why “just be present” can feel like an impossible demand.
